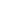









")
Pronunciation: DIP-luh-DOH-kuh-day
Author: Othniel Charles Marsh
Year: 1884
Meaning: Double beams (see etymology)
Locomotion: Quadrupedal (four legs)
Synonyms:
Atlantosauridae (Marsh, 1877)
Amphicoeliidae (Cope, 1878)
Dystrophaeidae (von Huene, 1904)
Apatosauridae (von Huene, 1927)
Atlantosauridae (Marsh, 1877)
Amphicoeliidae (Cope, 1878)
Dystrophaeidae (von Huene, 1904)
Apatosauridae (von Huene, 1927)
[Sereno, 1998]Definition
All diplodocoids more closely related to Diplodocus longus than Dicraeosaurus hansemanni.
About
Diplodocidae emerges in the Late Jurassic, appearing after fellow flagellicaudatans the dicraeosaurids but before the later-diverging rebbachisaurids. Their ancestry traces back to the Middle Jurassic diversification of diplodocoids, a time when semi-arid river systems and broad floodplains dotted with conifer and ginkgo stands influenced the herbivore guilds of the period. When diplodocids appear, they occupy landscapes where long reach and efficient foraging across extensive, seasonally dry habitats shaped their evolutionary trajectory.
Diplodocids are defined by a suite of elongations, reductions, and refinements that push the diplodocoid blueprint to its most extreme expression yet. Their skulls are elongated and narrow, with thin, pencil-like teeth restricted to the very front of the jaws for raking or branch stripping. Their forelimbs are markedly shorter than their hindlimbs, yet both are proportionally short compared to other sauropods, laying the foundation for an exceptionally long sweeping neck and equally long whip-ended tail. The vertebral column is extensively pneumatic, its internal structure reduced to a lattice of struts and rods that lightens the immense frame without sacrificing strength, a result of air-sac offshoots from the respiratory system invading the bones. High-set nasal openings perched between the eyes once prompted speculation about a trunk, but anatomical evidence shows that their skulls lacked the structural requirements for such soft-tissue elaboration. Within the family, two major subclades express these traits in different proportions, from the heavier-built apatosaurines to the more gracile, hyper-elongated diplodocines.
Diplodocids reach their peak in the Late Jurassic, emerging as major players in herbivore communities across the broad terrestrial landscapes of North America, East Africa, and western Europe. Their decline begins as the Jurassic gives way to the Cretaceous, a transition marked by climatic shifts, changing plant communities, and the rise of titanosaurian sauropods that would come to dominate later ecosystems. By the Early Cretaceous, diplodocids are gone, but their legacy endures in the public imagination as the archetypal long-bodied sauropods whose silhouettes define the grandeur of Jurassic giants, and in science through the first published reconstruction of an entire sauropod skeleton (Brontosaurus excelsus; Marsh, 1883), the first complete sauropod skull to be described (Diplodocus longus; Marsh, 1884), and the first mounted sauropod specimen (Apatosaurus ajax AMNH 460; Matthew, 1905).
Click here to view Dinochecker's A-Z list of Diplodocids.
Etymology
Diplodocidae is derived from the Greek "diploos" (double) and "dokos" (beam), and the Latin "-idae" (family). The name was coined for group anchor Diplodocus by O. C. Marsh, who assumed the whole family had a similarly forked chevron ("double beam") on the underside of each tail vertebra, though this arrangement is not present in all members, or even unique to diplodocids.
Colour
Tess Gallagher and colleagues' studies of fossilized skin from juvenile Diplodocus at the Mother's Day Quarry revealed first an unexpectedly diverse range of scale shapes (2021), and later the presence of melanosomes—microscopic pigment structures that, in living animals, produce colours and patterns (2025). Melanosomes have been identified in a wide array of fossils before—from feathered theropod dinosaurs to ancient amphibians—but this marked the first time they had been documented in a sauropod. Together, these finds challenged traditional views of sauropods as drab, elephant-like giants, instead pointing to vibrant, patterned skin that could have served roles in camouflage or social signaling, offering a rare glimpse into the biology and ecology of these massive dinosaurs.
Relationships
References
• Marsh OC (1883) "Principal characters of American Jurassic dinosaurs. Restoration of Brontosaurus". Geological Magazine, 10(9): 385-388. DOI: 10.1017/S0016756800166786.
• Marsh OC (1884) "Principal characters of American Jurassic dinosaurs. Part VII. Diplodocidae, a new family of the Sauropoda". American Journal of Science, 27: 161-167.
• Barrett PM and Upchurch P (1994) "Feeding mechanisms of Diplodocus". Gaia, 10: 195-204.
• Myhrvold N and Currie PJ (1997) "Supersonic sauropods? Tail dynamics in the diplodocids". Paleobiology, 23(4): 393-409. DOI: 10.1017/S0094837300019801.
• Upchurch P and Barrett PM (2000) "The evolution of sauropod feeding mechanism". In Sues (ed.) "Evolution of Herbivory in Terrestrial Vertebrates".
• Witmer LM (2001) "Nostril position in dinosaurs and other vertebrates and its significance for nasal function". Science, 293(5531): 850–853. DOI: 10.1126/science.1062681.
• Upchurch P, Barrett PM and Dodson P (2004) "Sauropoda". Page 259-322 in Weishampel, Dodson and Osmolska (eds.) "The Dinosauria: Second Edition".
• Carpenter K and Tidswell V (2005) "Thunder Lizards: The Sauropodomorph Dinosaurs".
• Knoll F, Galton PM and López-Antoñanzas R (2006) "Paleoneurological evidence against a proboscis in the sauropod dinosaur Diplodocus". Geobios, 39(2): 215–221. DOI: 10.1016/j.geobios.2004.11.005.
• Schwarz D, Frey E and Meyer CA (2007) "Pneumaticity and soft-tissue reconstructions in the neck of diplodocid and dicraeosaurid sauropods".  Acta Palaeontologica Polonica, 52(1): 167-188.
• Whitlock JA (2011) "Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses". PLoS ONE, 6(4): e18304. DOI: 10.1371/journal.pone.0018304.
• Young MT, Rayfield EJ, Holliday CM, Witmer LM, Button DJ, Upchurch P and Barrett PM (2012) "Cranial biomechanics of Diplodocus (Dinosauria, Sauropoda): testing hypotheses of feeding behaviour in an extinct megaherbivore". Naturwissenschaften, 99(8): 637–643. DOI: 10.1007/s00114-012-0944-y.
• Baron MG (2021) "Tactile tails: a new hypothesis for the function of the elongate tails of diplodocid sauropods". Historical Biology, 33(10): 2057–2066. DOI: 10.1080/08912963.2020.1769092.
• Gallagher T, Poole J and Schein JP (2021) "Evidence of integumentary scale diversity in the late Jurassic Sauropod Diplodocus sp. from the Mother’s Day Quarry, Montana". PeerJ, 9: e11202. DOI: 10.7717/peerj.11202.
• Conti S, Tschopp E, Mateus O, Zanoni A, Masarati P and Sala G (2022)
"Multibody analysis and soft tissue strength refute supersonic dinosaur tail". Scientific Reports, 12: 19245.
DOI: 10.1038/s41598-022-21633-2.
• Gallina PA, Apesteguía S, Carballido JL and Garderes JP (2022) "Southernmost Spiny Backs and Whiplash Tails: Flagellicaudatans from South America". Page 209–236 in Otero, Carballido and Pol (eds.) "South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution".
• Gallagher T, Folkes D, Pittman M, Kaye TG, Storrs GW and Schein JP (2025) "Fossilized melanosomes reveal colour patterning of a sauropod dinosaur".
Royal Society Open Science, 12(12): 251232.
DOI: 10.1098/rsos.251232.
Acta Palaeontologica Polonica, 52(1): 167-188.
• Whitlock JA (2011) "Inferences of Diplodocoid (Sauropoda: Dinosauria) Feeding Behavior from Snout Shape and Microwear Analyses". PLoS ONE, 6(4): e18304. DOI: 10.1371/journal.pone.0018304.
• Young MT, Rayfield EJ, Holliday CM, Witmer LM, Button DJ, Upchurch P and Barrett PM (2012) "Cranial biomechanics of Diplodocus (Dinosauria, Sauropoda): testing hypotheses of feeding behaviour in an extinct megaherbivore". Naturwissenschaften, 99(8): 637–643. DOI: 10.1007/s00114-012-0944-y.
• Baron MG (2021) "Tactile tails: a new hypothesis for the function of the elongate tails of diplodocid sauropods". Historical Biology, 33(10): 2057–2066. DOI: 10.1080/08912963.2020.1769092.
• Gallagher T, Poole J and Schein JP (2021) "Evidence of integumentary scale diversity in the late Jurassic Sauropod Diplodocus sp. from the Mother’s Day Quarry, Montana". PeerJ, 9: e11202. DOI: 10.7717/peerj.11202.
• Conti S, Tschopp E, Mateus O, Zanoni A, Masarati P and Sala G (2022)
"Multibody analysis and soft tissue strength refute supersonic dinosaur tail". Scientific Reports, 12: 19245.
DOI: 10.1038/s41598-022-21633-2.
• Gallina PA, Apesteguía S, Carballido JL and Garderes JP (2022) "Southernmost Spiny Backs and Whiplash Tails: Flagellicaudatans from South America". Page 209–236 in Otero, Carballido and Pol (eds.) "South American Sauropodomorph Dinosaurs: Record, Diversity and Evolution".
• Gallagher T, Folkes D, Pittman M, Kaye TG, Storrs GW and Schein JP (2025) "Fossilized melanosomes reveal colour patterning of a sauropod dinosaur".
Royal Society Open Science, 12(12): 251232.
DOI: 10.1098/rsos.251232.





